Ionophores - Mode of Action and Effects on
Milk Yield and Milk CompositionJ.J. Kennelly, L. Doepel, and K. Lien
Dept. of Agricultural, Food and Nutritional Science, University of Alberta, Edmonton, AB, Canada T6G 2P5
Take Home Messages
Introduction
The most common ionophores used in Alberta are Rumensin (monensin) and Bovatec (lasalocid). These have been used in the beef cattle industry for many years to increase feed efficiency and reduce the incidence of digestive disorders. Ionophores have also been used for several years in dairy heifers to prevent coccidiosis and improve body weight gain and feed efficiency. In lactating dairy cows monensin can be used at a level of 22 ppm for the control of coccidiosis. While coccidiosis generally is not a problem with mature dairy cattle, other benefits from the use of ionophores in dairy cattle diets can be expected. In early lactation, when cows are consuming high concentrate diets, ionophores would be expected to improve feed efficiency, increase the precursors of milk components thereby increasing milk yield, and reduce the incidence of lactic acidosis and ketosis. In late lactation, when cows are fed high forage diets, ionophores can improve daily body weight gain and feed efficiency. The effects of ionophores on rumen fermentation suggest an increase in important precursors for milk lactose and protein synthesis and ionophores may therefore have an important role in altering milk composition. This paper briefly reviews the effects of ionophores on metabolism and examines their effects on milk production and composition.
Mode of Action of Ionophores and Effects on Metabolism
Monensin and lasalocid are carboxylic polyether ionophore antibiotics that are produced by various strains of Streptomyces. The mode of action and effectiveness of ionophores have been presented in several reviews (Bergen and Bates, 1984; Goodrich et al., 1984; Schelling, 1984; Sprott et al., 1988; Spears, 1990; Corah, 1991). A brief summary of these are provided here to outline the effects of ionophores on nutrient supply. Ionophores function primarily in the rumen and improve the efficiency of production by influencing energy metabolism and nitrogen metabolism. As well, ionophores tend to reduce the incidence of lactic acidosis, bloat, and ketosis through their effects on rumen fermentation.
Ionophores exert their effects via their influence on rumen microbial populations and metabolism. Gram (-) bacteria tend to be resistant to ionophores. Gram (+) bacteria are more sensitive to ionophores due to the structure of their cell membranes. Ionophores alter the movement of ions across the cell membrane of gram (+) bacteria forcing the bacteria to expend energy to maintain intracellular pH and ion balance (Bagg, 1997). This energy expenditure reduces the bacteria's ability for growth and reproduction, leading to their eventual death. The resultant change in the ratio of gram (-) to gram (+) bacteria favours the production of propionate and succinate and reduces acetate, butyrate, hydrogen, ammonia, and lactic acid.
Feeding ionophores increases the metabolic energy values of feeds. This increase in energy efficiency is due to an increase of the molar proportion of propionate. Propionate is synthesized at the expense of acetate and butyrate, resulting in a decline in the acetate:propionate ratio in the rumen. The increase in propionate production is a result of ionophore selectivity for gram (+) bacterial populations. Methane production is also reduced as a result of feeding ionophores primarily through the inhibition of hydrogen and formate producing bacteria. Schelling (1984) reported that feeding of ionophores leads to a 4 to 31% reduction in methane production. These changes in rumen fermentation would be expected to improve animal production by increasing whole body glucose supply. Another benefit of increased propionate production is reduced levels of ketone bodies in early lactation cows. A reduction in clinical and subclinical ketosis as a result of ionophore supplementation has been demonstrated by several researchers (Sauer et al., 1989; Erasmus et al., 1993; Abe et al., 1994).
In addition to energy metabolism, ionophores also affect protein metabolism. A secondary effect of the increase in propionate production is a potential sparing effect on amino acids normally used for gluconeogenesis (Bergen and Bates, 1984). Ionophores have also been shown to have a protein-sparing effect in the rumen by decreasing rumen proteolysis and amino acid deamination, thereby increasing ruminal escape of dietary protein while decreasing the flow of microbial protein (Bergen and Bates, 1984; Schelling, 1984; Haïmoud et al., 1995). Consistent with these effects is a decrease in rumen ammonia production. The feeding of ionophores also results in an increase in apparent nitrogen digestibility, averaging 3.5 and 3.8 percentage units for monensin and lasalocid, respectively (Spears, 1990). This may possibly be attributed to a higher ratio of dietary to microbial protein entering the small intestine, and to a reduction in fecal endogenous nitrogen losses.
Effect of Ionophores on Milk Yield and Milk Components
Lactose Content and Milk Yield
Lactose is the single most important osmotic constituent of milk. An increase in lactose synthesis in the mammary gland is followed by an increase in water secretion which in turn influences milk volume. The lactose content in milk remains relatively constant at about 5%. In the cow, 85% of lactose carbon is derived from glucose (Annison, 1983). Propionate and amino acids are the major precursors for gluconeogenesis. The increased supply of these precursors brought about by the action of ionophores in the rumen would be expected to increase glucose availability which might be expected to increase lactose synthesis and milk production (Bauman and Elliot, 1983).
Studies with dairy cattle show considerable variation in the response of milk yield and lactose production to ionophores. Table 1 summarizes the results of 5 studies which evaluated the influence of ionophores on milk yield and composition. No effect on milk production or milk lactose yield was observed in early lactation Holsteins given diets consisting of approximately 50% forage and 50% concentrate with added lasalocid (Dye et al., 1988). In the study by Knowlton et al. (1996) multiparous cows in one group showed a slight increase in milk and lactose yield, while those in a second group exhibited a significant decline in milk and lactose yield when fed lasalocid. A group of primiparous cows in this same study had significantly higher milk and lactose yields when fed lasalocid compared to a control group. Weiss and Amiet (1990) studied the effect of lasalocid (340 mg/head/day) on milk production and composition in Holsteins in mid- to late lactation. Compared to the control animals, milk production of the lasalocid treated cattle increased by 0.9 kg/d with a diet consisting of 65% forage and 35% concentrate, but this was not significant. Conversely, Johnson et al. (1988) observed a non-significant decline in milk production of 0.8 kg/day with lasalocid (550 mg/day) in Jerseys and Holsteins fed 50% forage and 50% concentrate.
Erasmus et al. (1997), in a study with 60 lactating cows, reported that lasalocid had no effects on milk yield and composition but that dry matter intake decreased as the level of lasalocid increased (0, 10 ppm, 20 ppm). These results indicate that lasalocid may increase the efficiency of nutrient utilization. Christensen et al. (1994) also found that lasalocid caused a reduction in dry matter intake without affecting milk production thereby improving the efficiency of milk production.
Lynch et al. (1990) reported that Jersey, Jersey-Friesian and Friesian cattle on pasture that received monensin controlled release capsule (CRC) at 46 days post-calving had significantly higher milk yield than untreated control cows. Monensin CRC is a slow-release rumen capsule which releases approximately 335 mg monensin per day over 95 days (Duffield, 1997). During the 14 week treatment period, milk yield for the monensin treated cows averaged 14.3 kg/day while for the untreated group it averaged 13.2 kg/day (P < 0.01). Hayes et al. (1996) also found a milk production response to monensin administered as a rumen release capsule with cows on pasture. Cows receiving the capsule produced 0.41 L/day more milk than did controls. However, monensin treated (300 mg/day) Holstein-Friesen cows fed on pasture with supplemental pellets produced 67 L less milk over the first 58 days postpartum than their untreated counterparts (Abe et al., 1994). Milk production in Holsteins consuming 40% hay and 60% concentrate and given 200 mg monensin/day increased at a higher rate (34.9 to 39.4 kg) between 26 and 42 days postpartum than those not given monensin (37.0 to 38.7 kg) and had a slightly higher milk persistency (Kube et al. 1988).
Table 1. Change in milk yield and milk components in dairy cows with ionophore supplementation compared to controls.
|
# of Cows |
Breed |
Ionophore+ mg/head/d |
Forage: Conc. |
Days in Milk |
Milk Yield, kg/d |
Fat |
Protein |
Lactose |
Ref |
|
Percentage Units |
|||||||||
|
12a |
Holstein |
360 L |
44:56 |
7-100 |
0.2 |
-0.16* |
0.08* |
-0.03 |
21 |
|
16a |
Holstein |
111 L |
50:50 |
35-119 |
0.19 |
-0.11 |
0.02 |
0.04 |
9 |
|
16a |
Holstein |
220 L |
50:50 |
35-119 |
0.11 |
-0.10 |
-0.02 |
-0.06 |
9 |
|
16a |
Holstein |
333 L |
50:50 |
35-119 |
-0.34 |
-0.14 |
-0.05 |
0.11 |
9 |
|
90 |
Jersey/ Friesian |
320 M |
100:0 |
46-144 |
1.02b |
-0.39 |
-0.04 |
ND |
26 |
|
24 |
Holstein/ Jersey |
550 L |
50:50 |
90-160 |
-0.80 |
-0.83* |
-0.04 |
ND |
18 |
|
32 |
Holstein |
340 L |
65:35 |
170-268 |
0.9 |
-0.20 |
-0.10 |
ND |
36 |
+
M = monensin; L = lasalocid*Significantly different from control
a
Experiment run as a 4x4 latin square design with 21 d periods.b
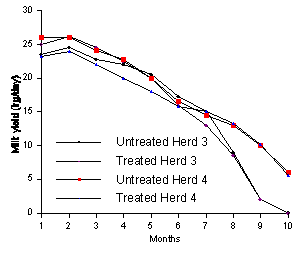
Daily milk yields are total milk yields averaged over number of days on experiment. Milk component concentrations calculated from total yields.Lean et al. (1994) completed an extensive study of the effects of monensin on milk production and composition involving 1061 cows in 6 different herds. The cows received monensin via a slow release intraruminal capsule and consumed varying amounts of forage and concentrates. The results from this study are consistent with other studies in the variability of the observed response to monensin. Overall, no significant differences were observed in milk yield in 5 of the 6 herds over the course of the lactation. In only one herd (Herd #2) was milk yield higher (P < 0.01) in monensin treated cows than control cows at all stages of lactation (Figure 1). In some of the herds, monensin treated cows had higher milk production than control cows for part of the lactation but then had lower production for the remainder of the lactation (Figures 1 and 2).
Figure 1. Milk yield for untreated and monensin-treated cows in herds 1 and 2
(adapted from Lean et al., 1994).Phipps et al. (1997) examined the effect of monensin on milk production in 98 cows over two consecutive lactations. Cows were given either 0 or 300 mg/d of monensin for 32 weeks in each lactation. In both lactations, milk yield was consistently higher for the monensin treated cows than for the control cows (Table 2). The increase in milk yield of the treated cows of 0.78 kg/d and 1.1 kg/d for lactations 1 and 2 respectively were not significant.
The difference in milk yield observations among the studies cited may be related to several factors - breed, level of concentrate in the diets, level of ionophore, time and length of ionophore treatment, and body condition of the animals. In
general, the observations from these studies suggest that the general usage of ionophores will have small but positive effects on milk yield but their use has also been reported to lead to detrimental effects on milk yield (Johnson et al., 1988; Abe et al., 1984; and Lean et al., 1994). Further studies are required to elucidate the conditions that promote the positive effects of ionophore supplementation.
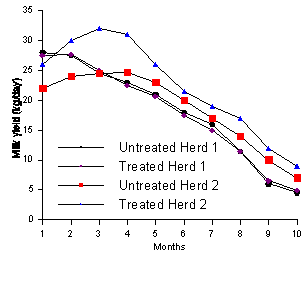
Figure 2. Milk yield for untreated and monensin-treated cows in herds 3 and 4. (Adapted from Lean et al., 1994).
Table 2. Mean milk production, adjusted by covariance for week prior to start of trial, when cows received either 0 or 300 mg/d of monensin.
|
monensin (mg/d) |
Lactation 1 |
Lactation 2 |
||||
|
0 |
300 |
SE1 |
0 |
300 |
SE1 |
|
|
Milk yield, kg/d |
21.4 |
22.2 |
0.38 |
27.2 |
28.3 |
0.72 |
|
Composition, g/kg |
||||||
|
Fat |
44.5 |
40.7 |
0.52 |
44.5 |
41.8 |
0.55 |
|
Protein |
36.4 |
34.8 |
0.22 |
35.5 |
34.4 |
0.32 |
|
Lactose |
46.7 |
46.3 |
0.19 |
46.6 |
46.2 |
0.17 |
|
Constituent yield, g/d |
||||||
|
Fat |
947 |
900 |
19.5 |
1195 |
1178 |
29.6 |
|
Protein |
775 |
776 |
13.6 |
949 |
973 |
21.5 |
|
Lactose |
1007 |
1035 |
19.5 |
1270 |
1308 |
34.5 |
1
Standard Error Least Square Means(Phipps et al., 1997)
Fat Content
Of the 3 major milk components, fat is the most amenable to dietary manipulation, both in terms of its concentration in milk and its composition. In early lactation, milk fat percentage exhibited a significant linear decline with increasing ionophore administration from 0 to 300 mg/head/day (Dye et al., 1988; Sauer et al., 1989). The latter authors noted a more severe milk fat depression at the lower level (15 g/t) of monensin (0.54 percentage units) compared to the higher level (30 g/t)(0.41 percentage units). In a study by Abe et al. (1994), milk fat was approximately 0.5 percentage units lower in monensin treated cattle at 30 and 60 days postpartum and more than 1.0 percentage unit lower at 90 days postpartum compared to control cattle. Duffield (1997) in his study of 25 herds found no significant treatment effect on milk fat percent. This is in contrast to the study by Phipps et al. (1997) in which the administration of monensin resulted in a large and significant drop in milk fat concentration (Table 2) which continued throughout the duration of the study (Figure 3).
In studies by Lynch et al. (1990) and Kube et al. (1988) there were only slight depressions in milk fat percentage with ionophore treatment. Johnson et al. (1988) reported that in mid-lactation fat percentage declined significantly by 0.58 percentage units in response to lasalocid treatment. It is interesting to note that in this trial the response from Jerseys was less than from Holsteins. Milk fat depression for Holsteins was 0.93 percentage units compared to 0.22 percentage units for Jerseys. Weiss and Amiet (1990) observed no milk fat depression in Holsteins in mid- to late-lactation with lasalocid. Knowlton et al. (1996) reported a significant decrease in milk fat percentage (0.16 percentage units) with 360 mg/head/day lasalocid As with milk yield data, the study by Lean et al. (1994) demonstrates the variability in milk fat yield response to monensin between herds and between stages of lactation within a herd. In general, there was no effect of monensin on milk fat yield.
The observed variability in the extent of milk fat depression associated with ionophore treatment may be due, at least in part, to the duration of the study. It appears that milk fat depression is more pronounced during the first weeks of ionophore addition. As the microbial population adjusts to ionophore inclusion, milk fat depression may be less pronounced.
A decrease in milk fat concentration, without a decline in milk yield, is not necessarily a negative consequence of using ionophores since there is interest in producing milk with a lower fat content (Kennelly, 1996). In addition, there is some evidence to suggest that the use of ionophores might promote a more favorable milk fatty acid composition. Kube et al. (1988) noted a significant increase in the concentrations of C16:1, C18:0, C18:1, C18:2, and C18:3 despite an overall depression in milk fat concentration with monensin supplementation.
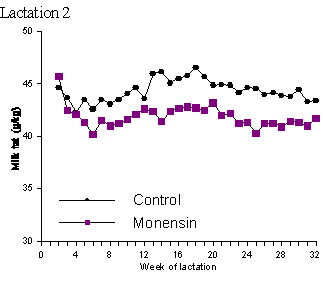
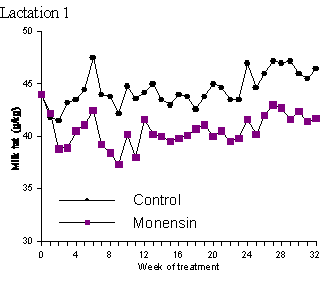
Figure 3. Mean milk fat concentration (g/kg) of control or monensin (300 mg/d) treated cows in lactation 1 and lactation 2 (adapted from Phipps et al., 1997).
Protein Content
In the vast majority of studies on ionophores and milk composition there has been no effect of ionophores on milk protein content (Dye et al., 1988; Johnson et al., 1988; Kube et al., 1988; Sauer et al., 1989; Weiss and Amiet, 1990; Abe et al., 1994; Duffield, 1997). In a study by Lynch et al. (1990) milk protein concentrations were significantly higher in a herd consisting of Jersey, Friesen and Jersey-Friesen cattle grazed entirely on pasture and given 320 mg monensin/day by rumen release compared to control animals. In Holsteins given diets consisting of approximately 50% forage and 50% concentrate with added lasalocid, milk protein content increased 0.08% compared to control groups (Knowlton et al., 1996). Lean et al. (1994) reported a slight increase (P = 0.1) in milk protein yield in monensin treated cattle in only 1 of 6 herds used in that study. One must be careful in interpreting milk protein data since alterations in the rate of degradation of dietary protein do not always increase true milk protein percentage, but may increase non-protein nitrogen at the expense of casein and/or whey protein (Khorasani et al., 1994; Robinson et al., 1991). Phipps et al. (1997) in a two-year study involving 98 cows given 0 or 300 mg/day of monensin for 32 weeks found that monensin treatment reduced milk protein content (Table 2; Figure 4). Milk from the treated cows had a protein content of 3.48% in the first lactation and 3.44% in the second lactation while milk from the control cows contained 3.64% and 3.55% protein in the 1st and 2nd lactations, respectively. As with fat, these studies indicate a need for a more detailed analysis of milk to better determine the effect of ionophores on milk composition
Mineral Content
The effects of ionophore supplementation on mineral availability and metabolism have been presented in recent reviews (Spears, 1990; Corah, 1991; Lean et al., 1997). Ionophore supplementation increases the apparent absorption of magnesium, phosphorus, zinc, selenium, and copper. The effects on calcium, potassium and sodium have been inconsistent. The potential exists for ionophores to alter the contents of some minerals in milk under certain conditions. Changes in the mineral content of milk as influenced by ionophores have not yet been reported. The ash content of milk is unaffected (Dye et al., 1988).
Conclusions
Additional data are required to elucidate the effects of ionophores on detailed milk composition. Studies to date are inconclusive in terms of the effects of ionophores on milk protein, fat and lactose. This may be due in part to experimental differences in breed, stage of lactation, forage:concentrate ratio, and level of ionophore supplementation. In addition to changes in milk yield, the potential benefits of ionophores include better feed conversion, accelerated weight gains, and reduced incidence of digestive disorders. Research needs to continue in the area of ionophore use in dairy cattle to determine the optimal level of supplementation both in terms of animal productivity and economic returns.
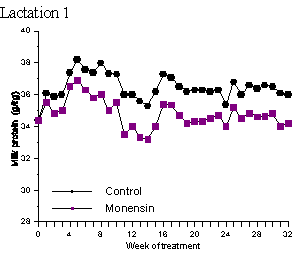
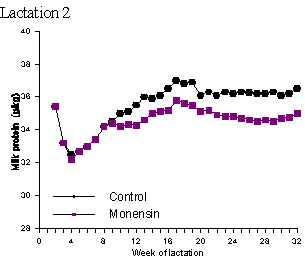
Figure 4. Mean milk protein concentration (g/kg) of control or monensin (300 mg/d) treated cows in lactation 1 and lactation 2 (adapted from Phipps et al., 1997.
References