Digestibility of Forage Fiber - Variation,
Measurement, and Relationship to Animal PerformanceMike Allen and Masahito Oba
Department of Animal Science, Michigan State University, East Lansing, MI 48824-1225 U.S.A.
Take Home Messages
Introduction
Variation in the digestibility of forage fiber affects the concentration of energy in the diet, the energy available for microbial protein synthesis in the rumen, and dry matter intake of ruminants. Therefore, it is an important factor affecting ruminant productivity. Yet, we do not routinely measure fiber digestibility or account for its variation when balancing diets for ruminants. This is primarily because fiber digestibility is poorly understood and difficult to predict by laboratory analyses. The digestion of fiber is a complex process that is affected not only by the forage, but also by the retention time of forage particles in the different segments of the gastrointestinal tract and by the amount and activity of enzymes secreted by fibrolytic organisms in the rumen and large intestine. The purpose of this paper is to assess what we know and don't know about fiber digestibility of forages including its measurement, prediction, variation, and relationship to animal performance.
Forage Fiber Fractions
Fiber is the fraction of plants that cannot be digested by mammalian enzymes. Forage fiber is composed of complex carbohydrates including cellulose, hemicellulose, and pectins, as well as lignin. Lignin is not a carbohydrate but a polyphenolic substance that is resistant to fermentative degradation and provides no nutritive value to the animal. Cellulose and hemicellulose are incompletely fermented to VFA's and extent of their fermentation is limited by the degree of lignification. Pectin is generally rapidly and completely fermented to VFA's. As forages mature, cellulose, hemicellulose and lignin contents increase and pectin content decreases. The hemicellulose content of forages ranges from 20 to 30% for grass and 8 to 12% for alfalfa while cellulose content ranges from 15 to 30% for both grasses and legumes. The lignin content of legumes is greater than for grasses at a similar maturity; lignin ranges from less than 6% to over 10% for legumes, less than 3% to over 7% for grasses, and from 2% to 4% for corn silage. Although pectin content of grasses are low, vegetative alfalfa may have levels that exceed 6%.
Neutral detergent fiber (NDF) is a measure of the cellulose, hemicellulose and lignin fractions of feeds; in doing so, it separates the highly digestible fractions of feeds with the less digestible, non-uniform fractions. Waldo (18) suggested that NDF is the best single chemical predictor of voluntary dry matter intake by ruminants. It is used in mechanistic models to predict energy content of forages (5,9) and dry matter intake for ruminants (1,9). It is also used as a measure of fiber requirements for dairy cattle (2). Neutral detergent fiber will be used as the measure of fiber with which to discuss fiber digestibility of forages in the remainder of this paper.
Variation in NDF Digestibility
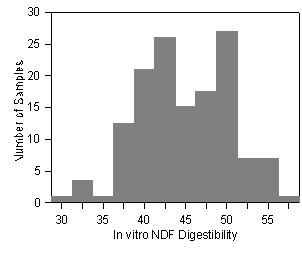
Ruminal digestibility of forage NDF has been reported to range from less than 35% to over 75% for forages (Table 1). The ranges reported were from experiments in the literature with no standardization for type of animal, level of intake, etc. Although fiber digestibility of forages is not constant for all animals and feeding conditions, much of this variation is due to the composition and structural differences of the forages. Removing animal and diet variation by using in vitro fermentation also resulted in a large variation in NDF digestibility for corn and alfalfa forages (Figure 1).
Not all NDF is potentially fermentable due to lignification. The indigestible fraction of forage NDF is a major factor affecting the utilization of fiber carbohydrates as it varies greatly and may exceed one half of the total NDF (Table 2). The ruminal digestibility of the fraction which is potentially fermentable is determined by how quickly it ferments (rate of digestion) and how quickly it passes from the rumen (19). Therefore, the rate at which the potentially fermentable NDF is fermented is another major factor affecting fiber carbohydrate utilization. Smith et al. (15) reported that rate of fermentation of forages measured in vitro ranged from less than 5% per hour to over 20% per hour (Table 2). While measurement in vitro with ground forages overestimates rate of fermentation of intact forages consumed by ruminants (see below), it does demonstrate the large differences among forages. It can be seen in Table 2 that as forages mature, the indigestible fraction of fiber increases and the rate of digestion of the potentially fermentable fraction decreases. Because of this, fiber digestibility generally decreases as forages mature within a cutting. However, environmental factors such as light intensity, day length, temperature, and soil moisture affect the relationship between fiber digestibility and maturity.
Table 1. Ruminal neutral-detergent fiber (NDF) digestibility of various forages. From Nocek and Russell (12).
|
Forage |
Ruminal NDF digestibility |
|
Alfalfa hay |
33-63 |
|
Alfalfa silage |
31-41 |
|
Corn silage |
32-68 |
|
Orchardgrass hay |
53-63 |
|
Orchardgrass silage |
41-48 |
|
Red clover hay |
31-59 |
|
Timothy hay |
66-77 |
|
Timothy silage |
49-52 |
Table 2. Rate of potentially digestible NDF digestion (% per hour) and indigestible NDF percent of various legumes and grasses at different maturities. From Smith et al., (15).
|
Forage |
Harvest Date |
Height (in) |
Maturity |
Indigestible NDF% |
Rate of NDF dig. (%/h) |
|
Alfalfa |
5/5 |
20 |
Pre-bloom |
45 |
19.1 |
|
6/16 |
45 |
Early pod |
58 |
7.3 |
|
|
Birdsfoot |
5/5 |
12 |
Pre-bloom |
51 |
17.3 |
|
Trefoil |
6/16 |
31 |
Early pod |
61 |
6.0 |
|
Bromegrass |
5/5 |
16 |
Vegetative |
15 |
18.3 |
|
6/16 |
55 |
Mature |
50 |
7.3 |
|
|
Orchardgrass |
5/15 |
28 |
Boot |
26 |
7.7 |
|
6/16 |
55 |
Mature |
46 |
4.6 |
1a. Corn Forage
1b. Alfalfa
Figure 1. Variation in digestibility of neutral detergent fiber (NDF) of (1a) corn forage samples (32 hybrids grown in 4 locations in 1988 and 1989) and (1b) alfalfa samples (10 cultivars grown in 2 locations, harvested with 4 cutting schedules for 4 cuttings each for 1990 and 1991). Allen et al. unpublished.
The third major factor affecting ruminal NDF digestibility is the rate at which the fiber passes from the rumen. Rate of passage of particulate matter in dairy cattle is affected primarily by level of intake, although many other factors such as feed particle size, dietary fiber level, and rate of digestion of the potentially digestible fiber fraction also affect passage rate. Until fairly recently, particle size reduction was thought to be the major limitation to passage from the rumen. However, as much as half of the particulate matter in the rumen is smaller than the largest particles in the feces indicating that something else must be involved in determining rate of passage. Particle buoyancy might be that other factor (17). Particles are buoyant when they are actively fermenting; gases (carbon dioxide and methane) produced during fermentation that are associated with the feed particles make them float in the rumen when they would otherwise sink. Buoyant particles might become suspended in the fibrous mat in the rumen and have a low probability of escape from the rumen. When the fermentable fiber content of a feed particle has been exhausted, the particle may sink to the bottom of the rumen and rumen movements will propel it towards the reticulum where it may escape the rumen through the reticulo-omasal orifice.
Both the indigestible fraction of NDF and the rate of digestion of the digestible fraction tends to be lower for grasses than legumes at the same maturity (Table 2). Because of this, grasses may have a longer retention time in the rumen than legumes because they may be buoyant for a longer period of time. As the fermentable fiber fraction of feed particles decreases, less gas is produced and particles may become less buoyant and sink. Particles which have low concentrations of fermentable fiber that ferments quickly, such as from alfalfa, might pass more quickly than particles which have more fermentable fiber which ferments slowly, such as from grasses. Figure 2 shows a comparison of the fermentation curves for dry matter and NDF of a typical grass and legume. As can be seen, NDF digestibility increases for both as retention time in the rumen increases. Rate of NDF fermentation is generally higher for legumes than for grasses at similar maturities (Table 2). Because of this, NDF digestibility at short ruminal retention times may be higher for legumes than for grasses (Figure 2).
Because the fraction of NDF which is indigestible is generally lower for grasses than for legumes (Table 2), grass NDF may have a higher digestibility at longer ruminal retention times. Longer ruminal retention times of grasses due to greater buoyancy over time will tend to increase the digestibility of grass NDF compared to legume NDF. Although grass NDF is generally more digestible than legume NDF, it may also be more filling and reduce intake because of an increased ruminal retention time. When intake is limited by ruminal fill of undigested feeds, legumes may allow higher intake than grasses as legume NDF ferments faster, and probably sinks and passes from the rumen faster than grass NDF.
This knowledge can be applied as a practical feeding strategy since rumen retention time of digesta decreases with level of intake. Assuming a rumen retention time for NDF of 30 hours for early lactation cows and 48 hours for late lactation cows, the potentially digestible NDF fraction of alfalfa may be nearly completely digested in the rumen of early lactation cows while that of grass is not. At lower ruminal retention times, legumes may have greater DM digestibility because of their lower NDF contents and similar NDF digestibility compared with grasses (Figure 2). Faster depletion of the potentially digestible fiber content of particles may allow faster passage and greater intake. On the other hand, grass may have greater NDF digestibility when fed to cows with longer ruminal retention times such as late lactation and dry cows. Grasses may have similar or greater DM digestibility than legumes when offered to cows with longer ruminal retention times. Because it isn't possible to harvest all forage with high fiber digestibility, if forages of different quality are stored separately, they can be allocated to animals that will benefit the most. Dry matter intake of cows in early lactation is limited by physical fill to the greatest extent and offering these animals fiber sources that digest and pass from the rumen more quickly may greatly increase their energy intake. On the other hand, fiber and dry matter digestibility of grasses will be greater when offered to cows with longer ruminal retention times such as late lactation and dry cows. Energy requirements of mid-lactation cows are relatively easy to meet and physical fill is less likely to limit DMI. Their longer ruminal retention times can be used to advantage by offering them fiber which may ferment more slowly but have a higher potential digestibility.
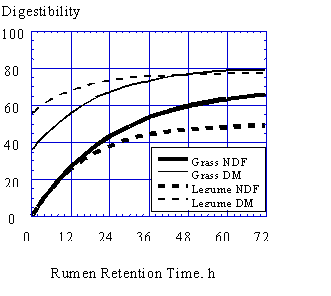
Figure 2. Fermentation curves for dry matter and neutral-detergent fiber of a typical grass and legume. Allen et al. unpublished.
Measurement of Fiber Digestibility
The most accurate data for fiber digestibility comes from animal digestibility experiments in which fiber intake and excretion in the feces is measured over time. This requires complete collection of fecal output for determination of weight and fiber content and is very labor intensive. Animal digestibility experiments using internal markers (indigestible ADF, indigestible NDF, acid insoluble ash, etc.) or external markers (chromic oxide, chromium mordanted fiber, rare earth elements, etc.) allow calculation of digestibility without complete fecal collections and have been more commonly implemented to measure digestibility of feed components with a variable loss in accuracy depending upon the technique used (16). Partitioning whole tract digestibility into ruminal and post-ruminal digestibility is commonly done using duodenally cannulated animals and one or more external markers assuming that little digestion occurs in omasum and abomasum because of lower microbial activity and relatively rapid transit through these organs.
For many purposes, the use of animals to measure fiber digestibility is not practical. Evaluation of corn silage hybrids or alfalfa varieties that require field, location and year replications may result in hundreds of samples to be compared and it is not feasible to conduct feeding trials to determine digestibility on this number of samples. Several different methods have been used to evaluate large numbers of samples for fiber digestibility including fermentation by ruminal microbes (in vitro batch and in situ) and by hydrolysis by fibrolytic enzymes. These methods have been extensively reviewed for the prediction of DM digestibility by Weiss (20) who concluded that the best method currently available for estimating digestibility is in vitro fermentation. However, all three methods have several significant problems in common for prediction of digestibility:
(1) Forages must be ground prior to fermentation. Grinding is necessary because the animal grinds the forage by chewing, increasing surface area available to enzymes. In addition, because a small sample is required, grinding allows a representative sample of the entire forage to be used. Because grinding increases surface area available to enzymes, rate of NDF digestion has been reported to increase as particle size of grass stem internodes decreases (4). However, forages are normally consumed as long or chopped particles which are gradually reduced in size by initial mastication and subsequent rumination. These particles have a slow initial rate of digestion in the rumen because of limited enzyme access. As the particles are reduced in size, surface area increases, increasing rate of digestion (1). Grinding the forage results an unrealistically high initial rate of fermentation.
(2) Retention time of forages in the rumen must be estimated. Passage of digesta from the rumen varies across animals of different species, level of intake, and physiological states (pregnant, lactating). An incubation time must be chosen which corresponds to the ruminal retention time for the animal of interest. However, there is very little data with which to make a decision for the retention time to use. Rate of passage data using markers isn't useful because no ideal external markers exist that flow with the feed fraction of interest and do not affect the passage characteristics of the feed labeled. While markers may provide some useful data for treatment differences in some cases, absolute rates are required to estimate digestibility of NDF. In general, rate of passage is overestimated by using markers as markers migrate and pass partly with the liquid fraction and(or) decrease the buoyancy of the labeled particles. Rate of passage of fiber particles has been measured in vivo by dividing indigestible NDF intake rate by the pool size of indigestible NDF in the rumen (7). This gives much more realistic rates of NDF passage but requires a single forage to be fed to measure its retention time which is unrealistic for dairy cattle. A much more serious problem is that ruminal retention times are influenced by the fermentation characteristics of the feed that affect buoyancy (see above), and it is not a fair comparison to use the same retention time for all forage samples. Using the same retention time for all forages will overestimate digestibility for some and underestimate digestibility for others. The same is true for particles within a forage sample that undoubtedly have quite different retention times.
(3) Fiber disappearance (solubilization) is generally measured which might not equate to fermentable energy for all forages. Solubilized phenolics or other unavailable substances will overestimate energy available to microbes and the animal. Measurement of fiber digestion as the production of gas has an advantage in that gas is produced from fermentation. However, many other problems are involved in relating the production of gas to fiber digestion by ruminal microbes which need to be addressed before this method should be used routinely.
The problem of different retention times for different animals of interest can be addressed by measuring fermentation over time and calculating a rate of digestion. This rate can be used with an appropriate rate of passage to calculate the digestibility of the potentially digestible fiber fraction (3). However, as mentioned above, the appropriate rate of passage is difficult to guess given our current knowledge and rate of fermentation is affected by particle size. It is unlikely that we will be able to accurately mimic the particle size reduction process during fermentation in vitro and use of ground forages results in overestimates of rate of fermentation. Calculation of fermentation rate also requires separation of the potentially digestible and indigestible fiber fractions. This increases the complexity of the mathematical model used, decreasing estimation accuracy.
In general, rates of fermentation determined in vitro and in situ overestimate actual rate, primarily because ground forages are used. When used with rates of passage from marker data, estimates of NDF digestibility that are within the wide range published in the literature from animal experiments are obtained. Although both rates are unrealistically high, when used together, they compensate for each other when digestibility is calculated, resulting in an estimate that seems to be reasonable. However, the size of the ruminal NDF pool predicted using these rates is unreasonably low indicating that both actual rates of digestion and passage are lower. Rate of digestion has been calculated using ruminal pools and influxes of digestible and indigestible NDF which leads to rates which are much lower than those obtained with ground forages (1). Rate of digestion determined in vitro and in situ may be a meaningful way to compare forages but actual rates may be quite different and rates of digestion so obtained should be used with caution.
Relationship with Lignin
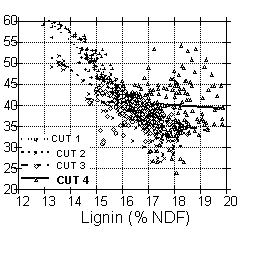
Lignin is the major component of fiber that limits digestion (10) and can be used as a crude indicator of NDF digestibility within a forage type. However, the relationship between lignin and NDF digestibility is not highly consistent within forages.
Figure 3. Variation in the relationship between lignin as a percent of NDF and 30 h in vitro NDF digestibility by cutting for 1,280 alfalfa samples (10 cultivars x 2 locations x 4 field replicates x 4 management schedules x 4 cuttings) grown in Michigan in 1990. Allen et al. unpublished.
It can be seen in Figure 3 that while there was a strong negative relationship between lignification of NDF and NDF digestibility for first cutting alfalfa, the relationship was not as strong for subsequent cuttings and no relationship was observed for the fourth cutting alfalfa data. Environmental factors also affect the relationship between lignin and fiber digestibility for corn silage. Figure 4 shows a negative relationship between lignification of NDF and NDF digestibility for corn forage grown in 1989. However, no relationship was observed between the two factors for the same corn hybrids grown in 1988 which was a drought year. Although lignification reduces NDF digestibility of forages, other factors may limit NDF digestion under some conditions.
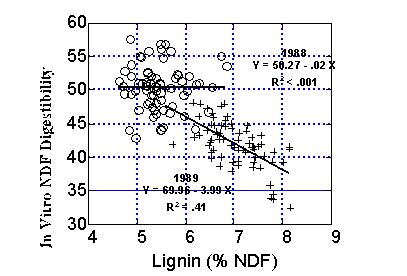
Figure 4. Relationship between lignin as a percent of NDF and 30 h in vitro NDF digestibility for corn forage (32 hybrids, 4 locations per year) grown in a dry year (1988) and a normal year (1989). Allen et al. unpublished.
Relationship with Animal Performance
The effect of fiber digestibility on animal performance is difficult to evaluate. Although it is thought that differences in fiber digestibility will result in greater energy intake, most experiments that have had treatment differences in fiber digestibility have been confounded by differences in NDF level, forage to concentrate ratio, and other dietary ingredients. Two of the most dramatic examples of animal response to forages that differed in fiber digestibility were obtained with growing lambs (11) and with mid-lactation dairy cows 8). Muller et al. (11) ensiled brown midrib and normal corn without ears and fed the silages to growing lambs. The NDF content of the two silages were the same but the lignin content of the brown midrib was 34% lower than the normal. The brown midrib silage was 10% more digestible than the normal with 20% greater NDF digestibility. The lambs offered the brown midrib silage had 29% greater dry matter intake than those offered the normal silage. Although the fiber contents of the two silages were the same, the fiber from the brown midrib silage was more digestible and it is presumed that it had less of a filling effect than the fiber from the normal silage. Grant et al. (8) offered a sorghum hybrid with the low-lignin BMR mutation and a different sorghum hybrid without the mutation in a total mixed ration (65% forage and 35% concentrate) to mid-lactation dairy cows. The BMR hybrid had higher NDF digestibility calculated from in situ and in vitro analyses and slightly higher NDF content. The BMR sorghum with higher NDF digestibility resulted in a 24% increase in DMI and a 46% increase in 4% fat corrected milk per day.
In a recent study, we evaluated the effect of enhanced NDF digestibility on DMI and milk production of high producing Holstein cows (13). At the beginning of experiment, cows were 89 ± 27.1 (mean ± SD) DIM, and produced 45.7 kg of milk per day. Experimental diets consisted of either brown midrib (bm3) corn silage or its isogenic normal counterpart (44.6% of DM). NDF digestibility estimated by 30h in vitro fermentation was higher for bm3 corn silage by 9.7 units. Diets were formulated for 19.6% CP, 31% NDF, and 56:44 FC ratio. DMI, milk production (3.5% FCM), SCM, and BCS were 2.0 kg (P<0.0001), 2.5 kg (P<0.0005), 2.6 kg (P<0.0001), and 0.09 units / 28d higher for the cows fed bm3 corn silage, respectively. No effect of treatment was observed for milk composition and body weight change. Enhanced in vitro NDF digestibility was associated with higher energy intake, which resulted in increased milk production and body condition. In addition, the highest producing cows had the greatest response in milk yield to the bm3 corn silage (Figure 5), presumably because these cows were limited by gut fill the greatest extent.
Although it is assumed that differences in NDF digestibility are at least partly responsible for the increased intake in the examples above, other differences among the treatments that affected intake might have existed. Another approach to evaluating the relationship of fiber digestibility and animal performance may be accomplished by statistical analysis of treatment means from the literature. We evaluated 19 sets of treatment means from 8 articles published in the Journal of Dairy Science for the effect of NDF digestibility on animal performance (14). Those articles which were used for this analysis had reported significant differences in NDF digestibility either in vivo, in situ, or in vitro. Data which compared fiber digestibility of legume with that of grass were not used because of the dramatic differences in diet NDF content or forage to concentrate ratio. NDF digestibility of forage significantly influenced DMI and milk production. One unit increase in NDF digestibility in vitro or in situ was associated with 0.15 kg increase in DMI and 0.26 kg increase in FCM (3.5%). Level of NDF concentration in a diet is negatively correlated to DMI since fiber ferments slowly and stays in the rumen longer than other feed components. Therefore, fiber that is more digestible might stimulate intake as it disappears from the rumen creating space for another meal sooner.
Improvement in Milk Yield (kg)
Figure 5. Relationship between initial milk yield and the milk yield response for cows fed bm3 corn silage with high NDF digestibility compared to isogenic normal corn silage. Data from Oba and Allen, unpublished.
Conclusions
Fiber digestibility of forages is variable and can affect dry matter intake, energy available to the animal, and energy available for microbial protein synthesis, and can have a great effect on milk yield. High producing cows or cows consuming high forage diets, that have intake limited by physical fill, will benefit the most from forages with high fiber digestibility.
References