Effective Fiber in Barley-Based Diets
Karen A. Beauchemin and Lyle M. Rode
Research Center, Agriculture and Agr-Food Canada, Box 3000, Lethbridge, AB T1J 4B1, Canada
E-mail: beauchemin@em.agr.ca
Take Home Messages
Introduction
To meet the energy requirements of lactating dairy cows, the proportion of concentrate in the diet is commonly increased, and high quality forages containing relatively low amounts of fiber are used. However, for dairy cows, diets that are low in fiber are associated with ruminal acidosis, reduced rumination, saliva secretion, and fiber digestion, lower acetate:propionate ratios, and milk fat depression. Coarse forage is needed in the diet to promote chewing and rumen motility and to maintain high concentrations of milk fat. Many of the recommendations for optimum fiber levels available to dairy producers in Western Canada were well developed for diets based on corn rather than barley grain. This presentation discusses the unique characteristics of barley versus corn, and how current fiber recommendations can be adapted for dairy cow diets based on barley.
Barley Digestion
Barley is the mainstay of the dairy cattle industry in western Canada. Digestible energy content is about 4% lower for barley than corn because barley contains more fiber and less starch. Barley is characterized by about 19 to 25% NDF compared with 9% for corn and the starch content of barley ranges from 52 to 73% compared to a range of 63 to 74% for corn (dry matter basis).
Grain Processing
Barley must be processed to maximize its availability in the rumen because whole barley kernels are not extensively damaged during eating (5). The rationale for processing is different for corn and barley. Because corn endosperm contains starch tightly packed within a protein matrix, the objective in corn processing is to release the starch from the matrix so that digestion can occur. In contrast, barley is encased within a hull which is very indigestible. However, barley starch is contained within an amorphous matrix within the endosperm and is digested within the rumen much more rapidly than corn starch. Therefore, processing of barley grain is designed to break open the hull so that the endosperm can be digested.
The easiest means to obtain maximum digestion is to finely grind barley. This completely exposes the starch to the microbes within the rumen. However, digestion occurs so rapidly that large quantities of finely ground barley will lead to acute acidosis and other metabolic disorders. Therefore, it is important to process barley sufficiently to break the hull without exposing too much starch for rapid digestion. This means maintaining a balance between over-processing (acidosis, metabolic diseases) and under-processing (reduced digestibility and animal performance).
Maintaining high ruminal digestion is more critical for barley than for corn. With corn, digestion that does not occur in the rumen is often compensated by digestion in the small intestine (Table 1). However, with barley, postruminal digestion is low if the grain is not extensively processed because the digesting enzymes in the intestine cannot penetrate the hull any better than the ruminal bacteria. This effect is demonstrated in Table 1 for hulless barley, that was only moderately processed (12). We observed low ruminal and post-ruminal digestion of starch. Because the hulless barley was under-processed in that study, much of the starch that was undigested in the rumen passed through to the feces, and total tract digestibility of starch was low.
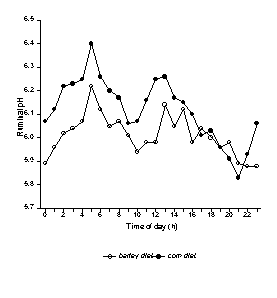
Because the extent of starch digestion within the rumen is greater for barley than corn, cows fed diets containing barley tend to have lower ruminal pH than those fed corn, when diets are formulated to contain the same amount of forage fiber (Figure 1). Although extensive processing of barley maximizes total tract digestion of feed, this also causes extremely rapid digestion of grain within the rumen causing dramatic swings in the degree of rumen acidity.
Table 1. Digestibility of starch by lactating dairy cows fed diets differing in grain source1 (12)
|
Item |
Diet |
SE |
||
|
Barley |
Hulless Barley |
Corn |
||
|
Dry matter intake, |
22.4 |
22.6 |
23 |
1 |
|
Starch intake, kg/d |
7.1 |
8.6 |
9 |
0.3 |
|
Starch flow, kg/d |
||||
|
Duodenal |
2.2c |
4.8b |
5.9a |
0.1 |
|
Fecal |
0.5b |
2.0a |
1.4ab |
0.3 |
|
Ruminal |
3.6 |
|||
|
Entering intestine |
3.8 |
|||
|
Total tract |
93.2a |
78.1b |
84.5ab |
2.4 |
1
Grains were steam-rolled.2
Diets consisted of 30% barley silage, 10% cubed alfalfa hay, and 60% concentrate.
Figure 1. Ruminal pH of dairy cows fed diets based on corn or barley grain. Both diets contained 22% NDF from forage sources and cows were fed at 0500 and 1300 h (12).
How Much Processing is Enough?
As rolled barley is processed finer, its bulk density (i.e. bushel weight) becomes less. By measuring the bushel weight after processing, as a percentage of the bushel weight before processing, we obtain a processing index (PI). In a survey of ten Alberta feed mills, we found that commercial feed mills tend to use steam-rolling to obtain a PI of 65 to 70% (10). This ensures that all grain is uniformly processed and no visible grain appears in the feces.
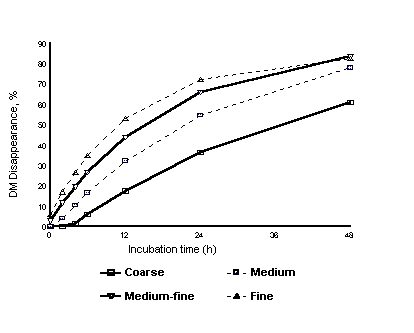
There is no one optimum degree of processing. We fed diets consisting of 30.5% barley silage, 8% alfalfa silage, 8% chopped alfalfa hay, 11% supplement, and 42.5% steam-rolled barley to dairy cows. The grain was rolled to a PI of 81% (coarse), 73% (medium), 64% (medium-fine), or 55% (fine). As the degree of processing increased, the rate and extent of ruminal digestion also increased, as expected (Figure 2). Increasing ruminal starch digestion by processing to a medium-fine degree resulted in the highest milk yield (Table 2). Processing the grain finer caused dry matter intake (DMI) to decline, likely due to high VFA production and lower ruminal pH. In this study diets were formulated to supply 32% total neutral detergent fiber (NDF) and 22% NDF from forage sources (NDF-f). Had a lower forage diet been used, the optimum PI would likely have been higher. In contrast, a diet containing more forage would allow the dairy producer to feed at a lower PI, thus gaining more digestible nutrients from the diet. Thus, the optimum amount of processing for barley depends on the amount of forage included in the diet.
Figure 2. Effects of processing steam-rolled barley on in sacco digestion kinetics measured in lactating dairy cows (Beauchemin, Rode, and Yang; unpublished data).
Table 2. Effects of degree of processing barley grain on dry matter intake and milk yield of dairy cows (Beauchemin, unpublished data).
|
Item |
Diet |
SE |
|||
|
Coarse |
Medium |
Medium Fine |
Fine |
||
|
Steam rolled barley |
|||||
|
Processing Index1 |
81 |
72.5 |
64 |
55.5 |
. . . |
|
Volume weight2, g/dL |
46.3 |
41.4 |
36.6 |
31.7 |
. . . |
|
Dry matter intake, kg/d |
18.7b |
21.4a |
21.7a |
20.1ab |
0.6 |
|
Milk yield, kg/d |
25.6c |
28.1b |
30.8a |
29.0b |
0.4 |
a,b
Corn and barley diets differ (P < 0.05).1
Processing index (PI) is measured as the bushel weight after processing, as a percentage of the bushel weight before processing.2
The volume weight of whole barley was 57.2 g/dL.Fiber Requirements
The dairy cow requires enough fiber to maintain an efficient and healthy digestive system (2). She has a requirement for both chemical fiber and physical fiber. Chemical fiber, measured as NDF, supplies fermentable carbohydrate that is required to support the microbial growth within the rumen. Physical fiber, measured as particle size, promotes chewing which results in the production of saliva. The buffers contained in saliva help minimize the risk of acidosis. The optimal amount of NDF in barley diets depends on the PI of the barley and the particle size of the forage.
Neutral Detergent Fiber (NDF)
Formulating diets for dietary NDF levels is a way of adjusting the forage-to-concentrate ratio of the diet to account for the variation in fiber content of forages. The NRC (9) recommends a minimum of 28% NDF in dairy cattle diets to maintain 3.5% milk fat and to minimize digestive disorders. It is also recommended that at least 75% of dietary NDF be supplied by forage. These recommendations are based on studies that mainly used corn rather than barley. Although this method of ration formulation is useful, NRC (9) guidelines are not directly applicable for barley-based diets. The NDF content of barley (19 to 25%) is higher than that of corn (7%), making it impossible to meet the NRC criteria for minimum fiber.
A commonly used thumb rule is to formulate the diet to supply 21% NDF from forage sources (NDF-f) without consideration of the NDF from concentrates (i.e., 28% NDF x 0.75 from forage = 21% NDF-f). To examine whether this recommendation is adequate for barley diets, we examined the relationship between NDF-f and milk yield and fat content (Figure 3). Treatment means from 5 experiments in which concentrates based on barley were fed to dairy cows are plotted in Figure 3 (3, 4, 6, 7, 12). Increasing the NDF-f content of the diet decreased milk yield, but increased milk fat content up to a maximum of 32% NDF-f. Beyond this level, milk fat content declined due to lower feed intake and insufficient energy intake. Therefore, within the range of forage fiber normally fed to lactating dairy cows, fat corrected milk was relatively unaffected by dietary NDF-f concentration.
The concentration of NDF-f necessary to maintain 3.5% milk fat in diets based on barley was extremely variable with the average ranging from 21 to 25%, which is higher than NRC (9) recommendations. This difference is attributed to the more extensive ruminal starch digestion of barley compared with corn (Figure 1). If the PI of the barley is low, and/or the particle size of the forage is fine, then the concentration of NDF-f necessary to avoid milk fat depression is at the higher end of the range. In contrast, if forage particle size is coarse and/or if the PI of barley is high, then less NDF-f is needed. Of course, for both barley and corn diets, the concentration of NDF-f necessary to prevent milk fat depression also depends on the genetics of the cows and other feeding practices, such as inclusion of high fiber byproduct feeds, frequency of feeding, and use of total mixed rations versus feeding grain and forage separately. If milk fat depression is not a primary concern, the minimum level of forage fiber necessary in dairy cow diets is that necessary to maintain healthy rumen function. That level is considerably lower than the level necessary to prevent milk fat depression. Thus, the optimum amount of NDF-f in the diet depends on the relative costs of concentrate and forage and the relative price of milk and fat.
Effective Fiber
The NDF assay assigns the same fiber value to long stemmed hay and ground hay, and does not predict the roughage value or effectiveness of the fiber. The physical components of fiber are often referred to as effective fiber, or effective NDF (eNDF). This pool of long fiber is chewed during eating and must be ruminated before it can leave the rumen. Saliva produced during chewing helps buffer acids produced in the rumen during microbial digestion of feed, thereby minimizing postprandial decreases in ruminal pH.
Figure 4 illustrates the relationship between the level of NDF-f consumed and the time spent eating, ruminating, and total chewing for diets consisting of barley concentrate and either long stemmed alfalfa hay, orchard grass hay, or corn silage (3). Time spent eating and ruminating increased as the amount of forage
Figure 3. Relationship between NDF from forage sources (NDF-f) and milk yield (top), 4% fat corrected milk (middle) and milk fat content (bottom) for dairy cows fed diets based on barley concentrates (3, 4, 6, 7, 12).
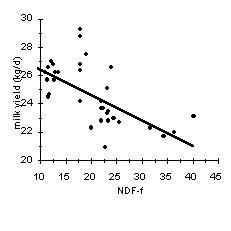
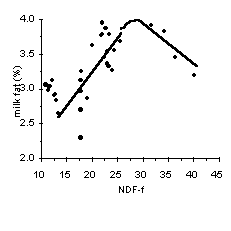
Figure 4. Relationship between the level of NDF from forage sources (NDF-f) consumed and the time spent eating (top), ruminating (middle), and total chewing (bottom) for diets based on barley concentrate (3).
fiber in the diet increased. The dairy cow typically spends 3 to 8 h/d eating and 6 to 9 h/d ruminating. The chewing time of cattle fed barley diets is considerable higher than those fed corn indicating a higher eNDF content of barley. In a study in which beef cattle fed 100% grain, rumination was twice as much when barley versus corn was fed (5). The higher rumination for barley is be due to the hulls, as well as a more resistant pericarp, resulting in a greater proportion of ingested intact kernels.
There is considerable difference among forages in the time spent chewing, even when compared at the same level of fiber intake (Figure 4). Eating time was much shorter for the diet containing corn silage due to the smaller particle size of corn silage compared with long hay. When feeds are chopped or ground, rumination and eating times are reduced (2). Grinding and pelleting feed results in a drastic decrease in chewing time, whereas the effect of chopping depends upon the fineness of chop.
Effective NDF measurements.
The Cornell Net Protein and Carbohydrate model (11) assigns eNDF values to individual feeds. For example, the eNDF content (% of NDF) of alfalfa hay is 92%, alfalfa silage is 82%, barley silage is 65%, and barley grain is 34% (Table 3). The values represent a combination of particle size measurements (i.e., measured as the proportion of NDF retained on a 1.18 mm square mesh screen after dry sieving) and subjective adjustments based on the feed digestion characteristics and their effects on ruminal pH. However, it is not clear how the eNDF values should be adjusted to account for on-farm variation in chop length of silage. Likewise, it is not specified how the adjustments for digestion characteristics are calculated.Table 3. Effective NDF (eNDF) content of silages estimated as the proportion of sample retained on the Penn State Particle Separator (PSPS) or from the Cornell model (11).
|
Silage type |
n |
top sieve1 |
bottom sieve2 |
eNDF |
|||||
|
Min |
Max |
Min |
Max |
PSPS |
Cornell |
||||
|
Heinrichs and Lammers 1997 |
|||||||||
|
Mixed |
1606 |
16 |
1 |
86 |
37 |
1 |
84 |
.. |
82 |
|
Grass |
582 |
18 |
1 |
77 |
41 |
5 |
83 |
.. |
.. |
|
Corn |
5395 |
8 |
1 |
82 |
51 |
0 |
89 |
.. |
81 |
|
Small |
529 |
15 |
1 |
62 |
48 |
12 |
90 |
.. |
65 |
|
TMR |
831 |
7 |
1 |
43 |
35 |
2 |
69 |
.. |
.. |
|
Beauchemin, unpublished data |
|||||||||
|
Barley |
57 |
10 |
2 |
42 |
37 |
15 |
54 |
63 |
65 |
1
Top sieve has 0.75-in circular openings.2
Bottom sieve has 0.31 in circular openings.An alternative is to use a commercially available sieving apparatus, such as the Penn State Particle Separator, to measure particle size. The Penn State Separator consists of a top sieve with 0.75-in circular openings and a bottom sieve with 0.31-in circular openings. Published results using the Penn State Separator are presented in Table 3. For legume and legume silage mixtures, an average of 16% or more of the particles (by weight) were retained on the top sieve, which was double that for corn silage. To maximize rumen function using the Penn State Separator, it is commonly recommended that 10 to 25% of the silage be retained on the top sieve (8). Legume silage stored in upright silos would likely be in the lower range (from 10 to 15%), whereas silages stored in bunker silos tend to have a greater proportion of long particles (from 15 to 25%). The middle sieve should have 30 to 40% of the material.
We used the Penn State Particle Separator to measure the eNDF content of barley silage from 57 farms in Alberta (Table 3). All silages were harvested at 3/8 in (i.e., 0.9 cm), but actual particle size distribution varied tremendously. In general, the proportion of long particles retained on the top sieve (10%) was low compared with other silages, and did not meet the target level of 10 to 25% on the top sieve. We calculated eNDF as the total proportion of sample retained by either sieve. The measured eNDF content of barley silage averaged 62.8% (±9.4), which is similar to the value of 65% used by the Cornell model. However, the values were extremely variable, ranging from 45.6 to 85.4%, with 30% of the samples having an eNDF content larger or smaller than the mean plus one standard deviation (Figure 5). Variations in particle size of barley silage must be accounted for when balancing rations for maintain healthy rumen function of cows.
Maximizing eNDF intake.
Particle size or eNDF content of forages is important, particularly if diets contain low levels of forage fiber. For example, we used alfalfa silage chopped at 5 or 10 cm (3/16 to 3/8") in barley diets containing less than adequate NDF-f (12%) or adequate NDF-f (22%) (4). The eNDF content of the fine and coarse silages was 47 and 77%, respectively, measured using the Penn State Particle Separator. For the low fiber diet, feeding the coarsely chopped silage increased rumination time, such that rumination time was similar to that of cows fed the higher fiber diet containing fine silage (Figure 6). Milk fat also increased when coarse versus fine silage was fed, but the extent of the increase was greater when high forage diets were fed. This is because increasing particle length of forages results in a larger intake of eNDF when forages comprise a larger portion of the diet.This study shows that increasing forage particle length in itself does not always overcome the milk fat depression associated with feeding low forage diets. In most cases, there is limited opportunity to increase silage chop length without compromising fermentation within the silo. Thus, it is much easier to increase eNDF intake by increasing the proportion of NDF-f rather than increasing particle length of silage. However, increasing the NDF-f content of the diet lowers the energy density of the diet and may decrease energy intake, resulting in lower milk yield. In that case, the extent of processing the barley should be considered.
Implications of Effective Fiber
The Cornell model (11) predicts ruminal pH from eNDF (% of DMI). However, the equation used is based on studies using dairy cows, sheep, and steers. To verify this relationship for lactating dairy cows, we compiled data for mean ruminal pH reported in 22 publications representing 95 diets fed to 619 cows. Ruminal pH was positively affected by level of NDF-f in the diet, but the variation in ruminal pH at a particular level of NDF-f was tremendous (Figure 7). Mean ruminal pH ranged from 5.6 to 6.7 within the NDF-f levels commonly fed to high producing dairy cows. This confirms that the minimum NDF-f level needed in the diet to minimize milk fat depression and ensure healthy rumen function is variable.
Using eNDF content (or eNDF intake) rather than NDF-f content did not improve the prediction of ruminal pH. It is possible that the eNDF values of feeds were not estimated correctly, thereby under or overestimating eNDF intake. However, it is likely that most of the variation is caused by differences in fermentability of the carbohydrate sources which is not accounted using the current system. This issue is extremely important if the model is to be used for diets containing barley concentrates because barley is extensively digested in the rumen causing low ruminal pH. Predictions of ruminal pH must account for the effective fiber content of the diet (saliva production) as well as the ruminal digestion of carbohydrates (acid production and absorption).
The Cornell model assumes that rumen pH is directly related to the efficiency of microbial yield (11). Microbial yield is reduced by 2.5% for each percentage unit reduction in eNDF < 20% of DMI, thus feeding low fiber diets increases the need for supplemental feed protein and ration costs. Accurate estimates of the efficiency of microbial protein synthesis are critical in predicting the amount of supplemental protein required and the cost of the diet. An example of this is given in Figure 8 where a diet was balanced for 48 kg of milk (3.5% fat; 3.12% protein) using the new Cornell Penn Miner (beta version) model. By increasing microbial protein synthesis by 50%, potential allowable milk yield increased from 48 to 92 kg/d in this theoretical application of the model. Clearly, the potential for altering the eNDF content of the diet to reduce the need for expensive protein supplements is large.
We do not have data for cows fed barley diets to verify the relationship between fiber and microbial protein synthesis for western Canadian dairy diets. However, we have shown that for diets containing the same proportion of forage (22% NDF-f), microbial protein synthesis is higher for barley diets than corn diets (12). Higher microbial protein synthesis was due to the greater proportion of ruminal fermented organic matter, rather than improved efficiency (Table 4).
Table 4. Digestibility of nitrogen by lactating dairy cows fed diets based on barley or corn grain (12)1
|
Item |
Grain |
SE |
|
|
Barley |
Corn |
||
|
Dry matter intake, kg/d |
22.4 |
23 |
1 |
|
Nitrogen intake, g/d |
558 |
562 |
27 |
|
Duodenal flow, kg/d |
|||
|
Total nitrogen |
546 |
526 |
27 |
|
Feed nitrogen |
240 |
268 |
19 |
|
Microbial nitrogen |
286a |
237b |
8 |
|
Efficiency of microbial |
23.4 |
24.4 |
3.1 |
|
Digestibility, % |
|||
|
Ruminal3 |
57.4 |
53.4 |
3 |
|
Postruminal |
72.3 |
71.8 |
0.5 |
|
Total tract |
73.5 |
74.2 |
1 |
a,b
Corn and barley diets differ (P < 0.05).1
Grains were steam-rolled and diets consisted of 30% barley silage, 10%cubed alfalfa hay, and 60% concentrate.2
Grams of microbial nitrogen / kilogram of organic matter truly fermented in the rumen.3
Corrected for microbial protein synthesis.Conclusions
The fiber required in barley-based diets to maintain high milk fat content depends upon the degree of processing the grain and the particle size of forage. Extensive processing of barley and fine-chopping of silage increases the need for forage fiber in the diet. If milk fat depression is not a concern, the minimum level of forage necessary in dairy cow diets is that necessary to promote healthy rumen function. That level is considerably lower than the level necessary to prevent milk fat depression. However, low-fiber diets may lower microbial protein synthesis and increase the need for supplemental feed protein.
References